

Ubiquitin-mediated degradation restricts spatiotemporal accumulation of the cytoplasmic male sterility protein WA352 to anthers in rice
Zixu Zhang, Zhi Ding, Xueye Feng, Jingjing Huang, Xu Peng, Yubin Xiao, Wubei Zong, Zhe Zhao, Yao-Guang Liu, Yongyao Xie, and Letian Chen
PNAS October 16, 2025; 122 (42) e2504381122; https://doi.org/10.1073/pnas.2504381122
Significance
Cytoplasmic male sterility (CMS) in plants helps breeders produce high-yield hybrid varieties. Genes that cause CMS are expressed throughout the plant, but the proteins they encode preferentially accumulate in anthers; how this occurs remains an unsolved mystery. Here, we show that mitochondrial F-box proteins participate in ubiquitination of the N-terminal transmembrane region of the CMS protein WA352, targeting it for degradation in vegetative tissues. However, the genes encoding these F-box proteins are specifically down-regulated at the microspore mother cell stage, allowing WA352 to accumulate and hijack the cytochrome c oxidase subunit COX11, causing a burst of reactive oxygen species in the anther and consequent CMS. These findings reveal a mechanism regulating the specific accumulation of this CMS protein in mitochondria.
Abstract
Cytoplasmic male sterility (CMS) is caused by mitochondrial genes that are constitutively expressed in plant tissues, although the encoded proteins preferentially accumulate in anthers. The mechanisms regulating CMS protein accumulation remain unclear. Here, we explored this process using wild-abortive CMS (CMS-WA) rice (Oryza sativa). We show that WA352, the causal protein of CMS-WA, is degraded by the ubiquitin–proteasome system (UPS). Structural analysis and protein truncation assays revealed that the N terminus of WA352 is critical for its anchoring to the inner mitochondrial membrane and its UPS-mediated degradation. Functional complementation confirmed that WA352151–352, lacking the N-terminal domain, accumulates constitutively in vegetative tissues, causing a reactive oxygen species burst and retarding rice growth. We further identified three mitochondrion-localized F-box proteins that participate in WA352 ubiquitination and degradation. Our findings demonstrate that UPS-mediated regulation restricts WA352 accumulation to anthers, allowing it to specifically disrupt anther development, thus helping to explain the male-specific effects of CMS genes in plants.
See: https://www.pnas.org/doi/10.1073/pnas.2504381122
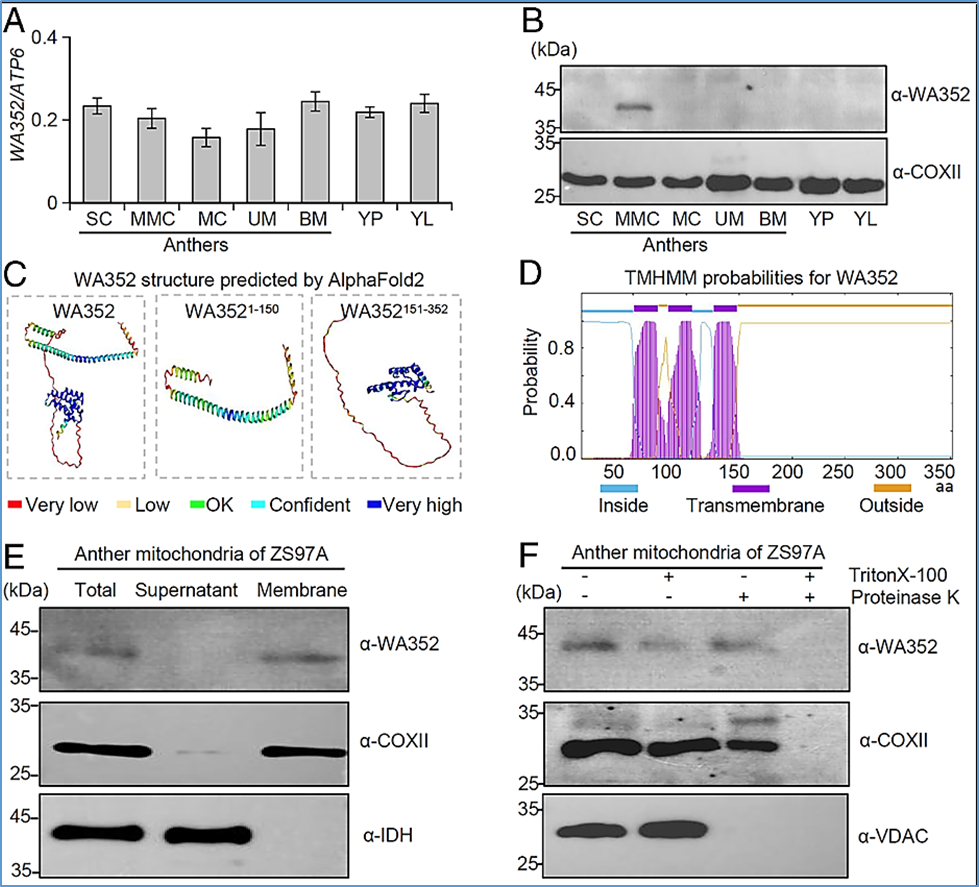
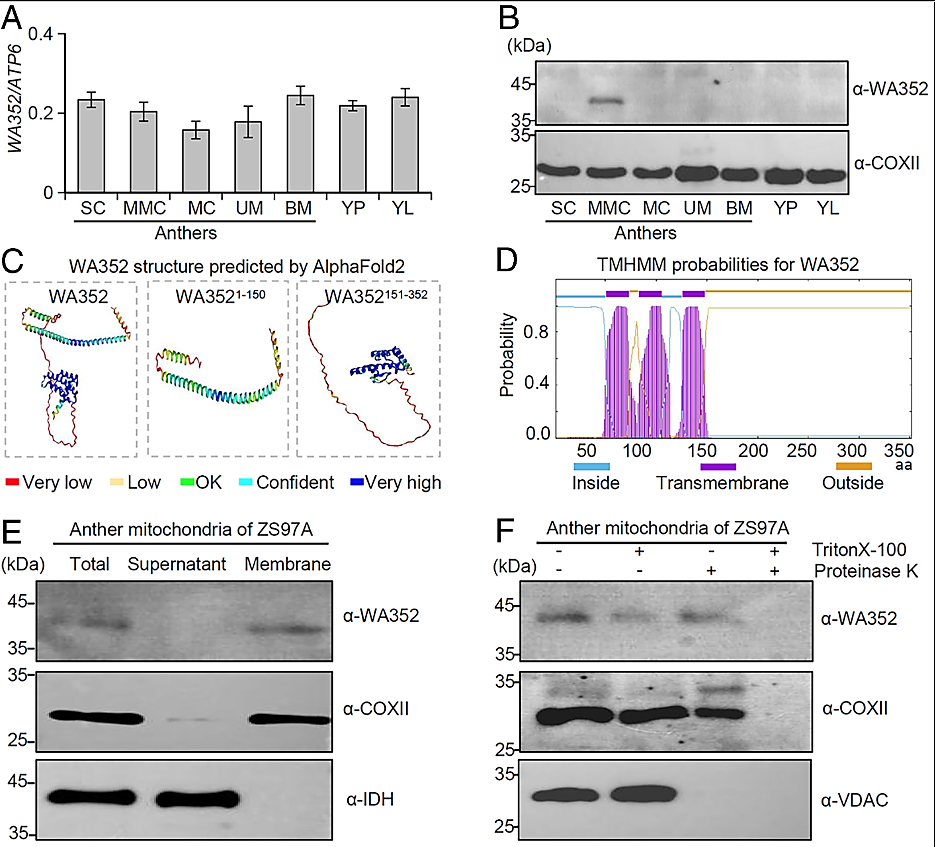 Figure 1:
Figure 1:
Expression profiling and structural analysis of the CMS protein WA352.
(A) The mitochondrial gene WA352 is constitutively expressed in anthers during the SC stage, MMC stage, MC stage, UM stage, BM stage, and in YP and YL of a CMS-WA line ZS97A. The mitochondrial gene ATP6 was used as an internal control, and WA352 transcript levels are shown relative to ATP6. Data are shown as means ± SD (n = 3). (B) Immunoblot of WA352 protein in different tissues and anther developmental stages of ZS97A. The mitochondrial protein COXⅡ was used as an internal control for loading, and stages are the same as in (A). (C) Structure of full-length and truncated forms of WA352 as predicted by AlphaFold2, showing an N-terminal transmembrane domain (amino acids 1 to 150, WA3521–150) and C-terminal conserved region (amino acids 151 to 352, WA352151–352). Amino acids are colored based on their per-residue confidence scores. (D) WA352 transmembrane domains predicted using TMHMM2.0. The three-transmembrane helical domain at the N terminus is shown in purple. (E) Mitochondria protein samples from anthers at the MMC stage were separated into soluble supernatant and membrane fractions and used to analyze the localization of WA352 by immunoblotting. COXⅡ was used as a control mitochondrial membrane protein, and IDH was used as a control soluble matrix protein. (F) WA352 localizes to the inner mitochondrial membrane. Mitochondrial fractions treated with (+) or without (−) the membrane-dissolving reagent Triton X-100 and the protein-digesting enzyme proteinase K. COXⅡ was used as an inner membrane protein control, and VDAC was used as an outer membrane protein control.
Views: 238