

Structural insights into ABA receptor agonists reveal critical features to optimize and design a broad-spectrum ABA signaling activator
Mar Bono, Cristian Mayordomo, Alberto Coego, Jonatan Illescas-Miranda, Maria Rivera-Moreno, Lourdes Infantes, Pablo López-Carracedo, Mayra Sanchez-Olvera, Constanza Martin-Vasquez, Gaston A. Pizzio, Javier Merino, Javier Forment, Ebe Merilo, Juan Carlos Estevez, Armando Albert, Pedro L. Rodriguez
MOLECULAR PLANT; Volume 18, Issue 9: p1526-1548; September 01, 2025
Abstract
Crop yield is at increasing risk due to water scarcity and climate change. Agrochemicals can activate hormone receptors to regulate transpiration and modulate transcription and address water deficits. Structure-guided optimization of multiple abscisic acid (ABA) receptor–agonist interactions is necessary to activate the entire PYRABACTIN RESISTANCE 1 (PYR1)/PYR1-LIKE (PYL)/REGULATORY COMPONENTS OF ABA RECEPTORS (RCAR) receptor family. The new agonist iCB, produced through scaffold-merging led by X-ray structure, activates subfamilies II and III at low-nM concentrations and subfamily I receptors at higher-nM concentrations. Structural analysis of opabactin and iCB ternary complexes reveals selectivity-determining residues, making the PYL1/PYL4/PYL8 subfamilies sensitive to specific agonists and highlighting the differential sensitivity of receptor subfamilies to agonists across plant species. iCB may activate most eudicots’ PYL8-like receptors, in contrast to opabactin, due to limited steric constraints. This enables iCB to activate PYL8-like receptors with a bulkier Leu residue in the 3′ tunnel, such as AtPYL8, SlPYL8, and VviPYL8. In contrast, opabactin activation is limited to receptors with Val at this position, for example, TaPYL8. Therefore, iCB extends its action to more ABA receptors than CB, iSB09, and opabactin, exhibits higher affinity than ABA for dimeric receptors, and can protect tomato plants against drought. In addition to regulating stomatal conductance and lowering water consumption, iCB protects photosystem II and improves photosynthesis following prolonged water deficit. Moreover, iCB induces an ABA-like transcriptional response, upregulates the osmolyte synthesis, and can be hyperpotentiated when combined with the expression of a customized receptor. Our results provide structural insights for optimizing agonist design and aiding plants in managing water deficits.
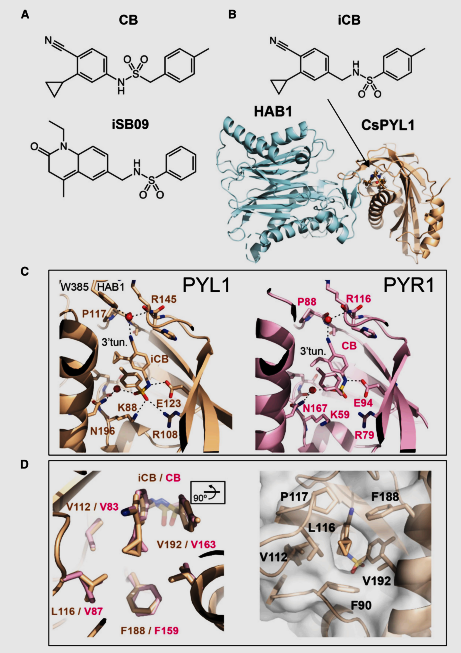
Figure 1: The structure and coordination of iCB in the ternary complex with CsPYL1 and HAB1. (A) Chemical structures of CB and iSB09. (B) Chemical structure of iCB. A ribbon representation of the CsPYL1(wheat)–iCB–HAB1(cyan) ternary complex. The iCB molecule in the ABAbinding pocket of CsPYL1 is highlighted in a balland-stick representation. (C) Structural analysis of iCB and CB (PDB: 5YR6) (Vaidya et al., 2017) in the ligand-binding pocket of CsPYL1 (wheat) and AtPYR1 (pink) receptors, respectively. For iCB, the Trp lock, which involves Trp385 from HAB1, is indicated. The figure illustrates critical differences between the binding patterns of iCB and CB. Dashed lines represent hydrogen bonds from 2.6 to 3.4 A˚ . (D) Hydrophobic interactions of iCB and CB in the 3′ tunnel of CsPYL1 and AtPYR1 receptors, respectively (left). Detail of the iCB’s cyclopropyl group in a surface representation of the hydrophobic 3′ tunnel of CsPYL1 (right), observed from a perspective rotated around 90◦ compared to what is depicted in the left panel.
Views: 139