

Editing strigolactone hormone receptor for robust antiviral silencing in rice
Guoyi Yang, Ming Wu, Shuai Zhang, Yucen Huang, Yixiao Liu, Xiyuan Yu, Jiayang Hu, Le Mi, Peng Gan, Yuansheng Wu, Jing Zou, Baogang Zhang, Qun Hu, Jie Hu, Ruifeng Yao, Bojian Zhong, Xianbo Huang, Huiting Xie, Yinghua Ji, Yi Li, Jie Zhang, Liming Yan, Shou-Wei Ding, Shanshan Zhao, Jianguo Wu
Cell; 2026 Feb 24: S0092-8674(26)00100-5. doi: 10.1016/j.cell.2026.01.013.
Abstract
The small interfering RNA (siRNA) pathway directs broad-spectrum antiviral defense through RNA silencing so that virulent infection requires efficient suppression of the defense mechanism. Here, we show that strigolactone (SL) hormone signaling promotes antiviral silencing in rice plants by transcriptional activation of RNA-dependent RNA polymerase 1 (RDR1) and RDR6. We demonstrate that protein P3 of the rice grassy stunt virus (RGSV) blocks SL signaling by directly sequestering the receptor DWARF14 from DWARF3. Structural and functional analyses of the P3-DWARF14 complex reveal that the aspartic acid at position 102 (D102) of DWARF14 is essential for the P3 interaction but not for SL perception. Notably, a single D102N substitution of DWARF14, introduced into two rice cultivars by cytosine base editing (CBE) confers resistance against RGSV by blocking viral suppression of SL signaling-dependent antiviral silencing. Our findings establish a transgene-free strategy for engineering disease resistance by precise genome editing of the SL receptor to escape pathogen suppression of the endogenous defense pathway.
See https://pubmed.ncbi.nlm.nih.gov/41742412/
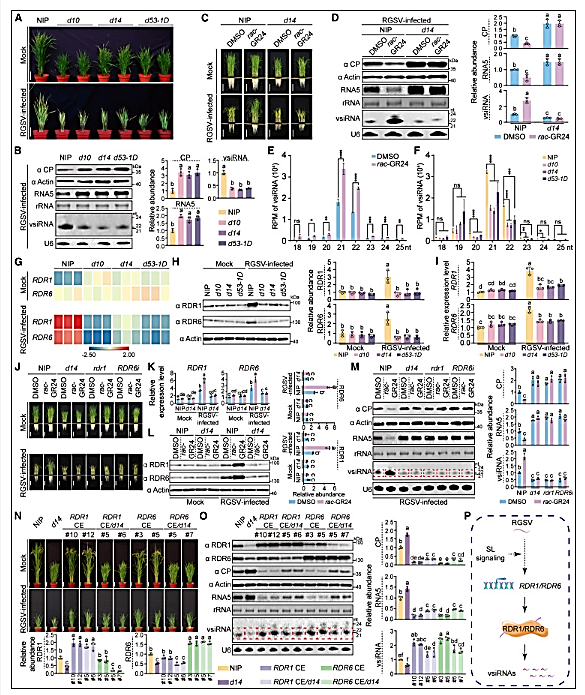
Figure 1: SL signaling enhances antiviral RNAi by promoting virus-induced activation of RDR1 and RDR6
(A) Photographs of mock-inoculated or RGSV-infected NIP and the indicated plants at 8 weeks post-inoculation (wpi). Scale bar, 10 cm.
(B and D) Immunoblot and northern blot analyses of RGSV CP, viral RNA, and vsiRNAs in the indicated plants.
(C) Photograph of mock-inoculated or RGSV-infected NIP and d14 rice plants with and without rac-GR24 treatment at 4 wpi. Scale bar, 10 cm.
(E and F) vsiRNA contents in RGSV-infected, rac-GR24-treated and -untreated NIP (E) and the indicated RGSV-infected plants (F), derived from small RNA-seq data.
(G) Heatmap indicating the relative expression of RDR1 and RDR6 in mock-inoculated or RGSV-infected NIP and the indicated plants.
(H and I) Immunoblot (H) and real-time qPCR (I) analyses of RDR1 and RDR6 levels in mock-inoculated or RGSV-infected NIP and the indicated plants at 4 wpi.
(J) Photographs of mock-inoculated or RGSV-infected NIP, d14, rdr1, and RDR6i rice plants with or without rac-GR24 treatment at 4 wpi. Scale bar, 10 cm.
(K and L) Real-time qPCR (K) and immunoblot (L) analyses of RDR1 and RDR6 levels in mock-inoculated or RGSV-infected NIP or d14 rice plants with or without rac-GR24 treatment.
(M and O) Immunoblot and northern blot analyses of RDR1, RDR6, RGSV CP, viral RNA, and vsiRNAs in the indicated plants.
(N) Photographs of mock-inoculated or RGSV-infected NIP and the indicated plants at 8 wpi. Scale bar, 10 cm.
(P) A summary model illustrating the regulatory role of SL signaling in the virus-induced RNAi response.
Data represent mean ± SD (n = 3). Statistical significance was determined by one-way ANOVA with Tukey’s test for (B) and (O); two-way ANOVA with Tukey’s test for (D), (H), (I), (K), (L), and (M). Two-tailed Student’s t test for (E) and (F): ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001, ∗∗∗∗p < 0.0001. ns, not significant.
Views: 152