

A yeast mating platform for multiplex screening of fungal GPCR–ligand interactions
Giovanni Schiesaro, Melani Mariscal, Mathias Jönsson, Ricardo Tenente, Mathies Brinks Sørensen, Marcus Wäneskog, María Victoria Aguilar-Pontes, Agustina Undabarrena, Marcus Deichmann, Emma E. Hoch-Schneider, Viji Kandasamy, Thomas M. Frimurer, Antonio Di Pietro, Line Katrine Harder Clemmensen, Michael Krogh Jensen, and Emil Damgaard Jensen
PNAS; October 24, 2025; 122 (43) e2521198122; https://doi.org/10.1073/pnas.2521198122
Significance
Fungal pathogens rely on G protein–coupled receptors (GPCRs) to sense environmental cues and coordinate host infection. By establishing a yeast mating platform for multiplex GPCR–ligand screening, we identify agonist and antagonist peptides that can interfere with fungal cell–cell communication. This work not only accelerates the study of fungal GPCR–ligand interactions but also demonstrates, for the phytopathogen Fusarium oxysporum, that interfering with GPCR-mediated cell–cell communication is a promising target for antifungal strategies in agriculture.
Abstract
Fungi are essential members across ecosystems, yet phytopathogenic fungi pose an increasing risk to crop yields. Despite their ecologic importance, cell–cell communication in fungi is underexplored, partly due to the lack of high-throughput techniques. Here, we developed a Yeast Mating Platform (YeMaP) to investigate the interaction between fungal G protein–coupled receptors (GPCRs) and pheromone peptides. We used YeMaP for high-throughput screening of 8,000 pheromone sequences and identified peptides with improved agonism or antagonism action. We found that these peptides can be applied in a native fungal system such as the plant pathogen Fusarium oxysporum, to control hyphal chemotropism and reduce plant root penetration. Additionally, we utilized YeMaP in a one-pot assay to investigate how abiotic factors influence the communication of multiple pheromone–GPCR combinations and found that the cell–cell communication mediated by the GPCR Ste2 from F. oxysporum signaled robustly across different abiotic factors, while other fungal GPCR–pheromone interactions were more sensitive to changes. Taken together, YeMaP accelerates the identification of fungal GPCR–peptide interactions by enabling one-pot assays, and serves as a model system for studying fungal cell–cell communication.
See https://www.pnas.org/doi/10.1073/pnas.2521198122
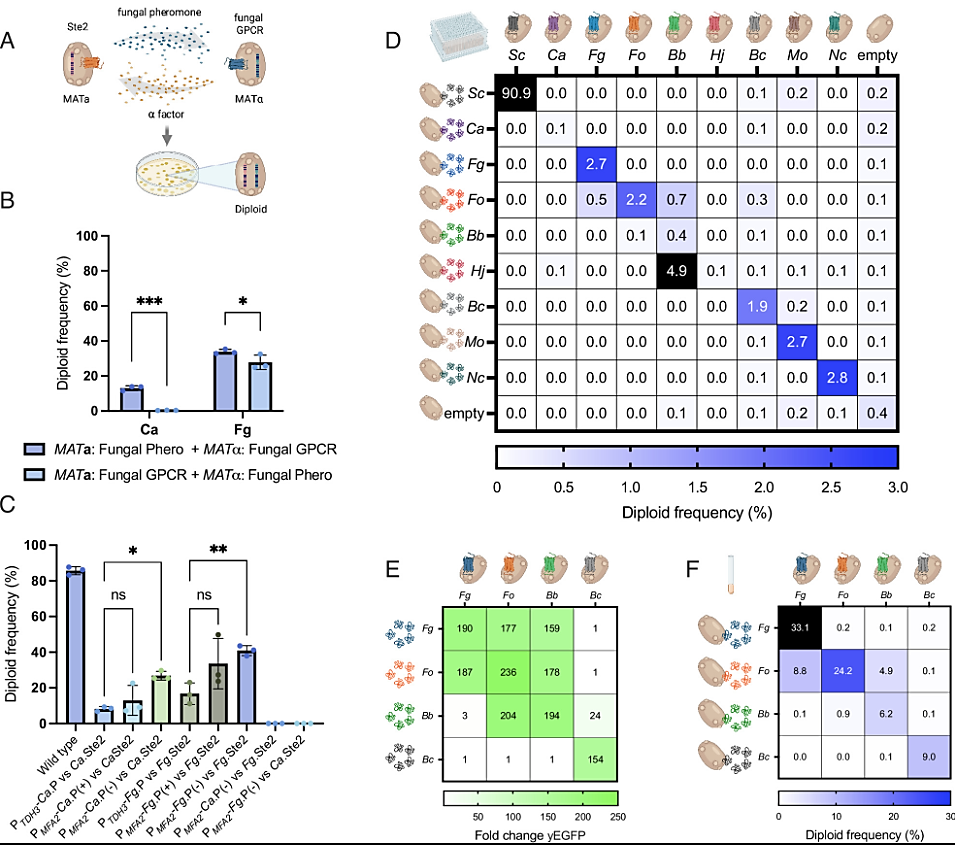
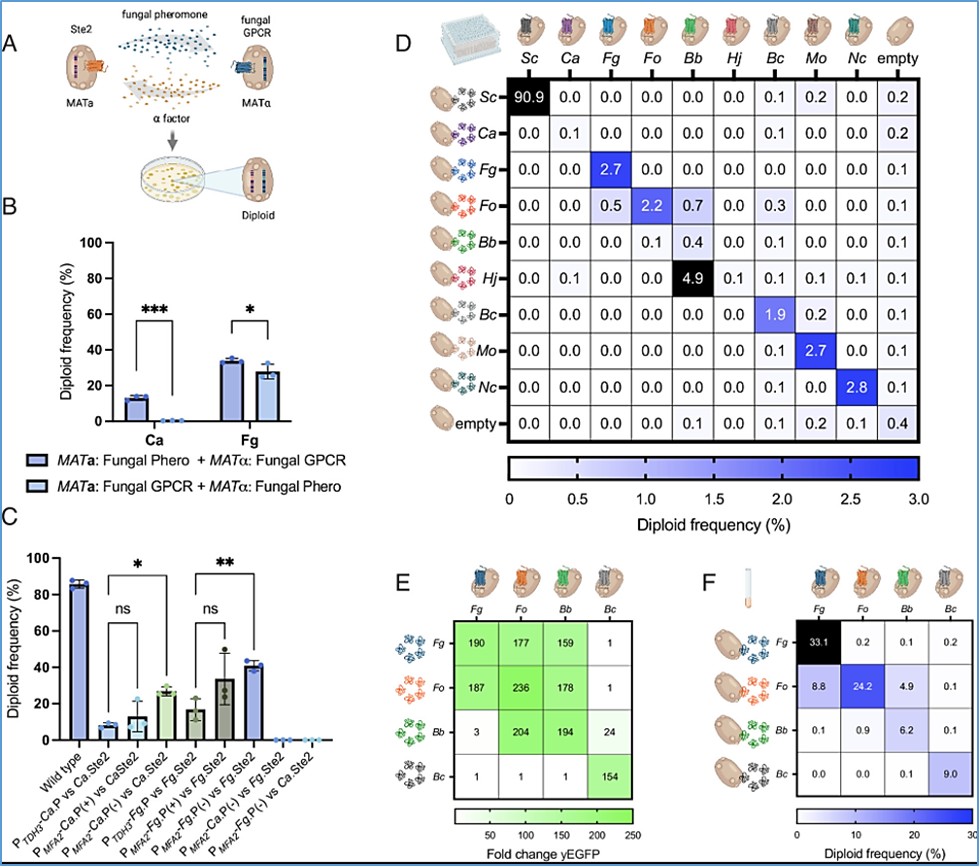
Figure 1
YeMaP correctly identifies fungal GPCR–pheromone interactions.
(A) The One-on-One setting involves coculturing two yeast strains in which half of the mating communication system has been replaced with heterologous fungal components; one cell expresses a given fungal Ste2 GPCR homolog while a second cell secretes a given fungal alpha pheromone. After successful mating, diploid cells are formed and selected. (B) Comparison of cocultures expressing C. albicans (Ca) or F. graminearum (Fg) components under the following conditions: MATa cells expressing either the Ca or Fg alpha pheromone from the MFA2 promoter (Ca: GEN29, Fg: GEN35) and MATα cells expressing either the Ca or Fg Ste2 GPCR (Ca: GEN36, Fg: GEN37); or MATa cells expressing the fungal GPCR (Ca: GEN100, Fg: GEN78) and MATα cells expressing the Ca or Fg alpha pheromone from the MFα1 promoter (Ca: GEN76, Fg: GEN77). MFA2 and MFα1 promoters were selected as the strongest inducible pheromone promoters based on previous studies (20, 25). (C) Diploid frequency comparison between different designs. From left to right: GEN18 and GEN27 were cocultured as a positive control (Wild type); GEN36 (Ca.Ste2) cocultured with GEN28 (PTDH3-Ca.P); with GEN29 (PMFA2-Ca.P); or with GEN55 (PMFA2-Ca.P, mfa1Δ, mfa2Δ). GEN37 (Fg.Ste2) cocultured with GEN34 (PTDH3-Fg.P); with GEN35 (PMFA2-Fg.P); or with GEN58 (PMFA2-Fg.P, mfa1Δ, mfa2Δ); and GPCRs combined with noncognate pheromone GEN55 + GEN37; and GEN58 + GEN36. (D) Yeast mating matrix with MATα strains individually expressing eight different fungal Ste2 GPCRs (columns), cocultured with either of eight alpha pheromone-secreting MATa strains (rows). As positive and negative controls, respectively, strains expressing either Sc Ste2 or alpha pheromone, or strains lacking a GPCR or a pheromone were used. Blue gradient is set to a maximum of 3% of diploid frequency, with values >3% colored in black. Complete list of organisms and pheromone sequences is in SI Appendix, Table S1. (E) Activation levels of Fg.Ste2 (CPK366), Fo.Ste2 (GEN70), Bb.Ste2 (GEN71), and Bc.Ste2 (GEN128) biosensors (columns), incubated with 10 μM alpha pheromone from Fg, Fo, Bb, or Bc (rows). The fluorescent signal was normalized to the yEGFP background value (receptor without pheromone). (F) Validation of the mating matrix in culture tubes. Strains individually expressing the four indicated GPCRs (columns) were cocultured with those secreting the indicated alpha pheromone (rows). Blue gradient is set to a maximum of 30% of diploid frequency, with values >30% colored in black. Plating was performed in three technical triplicates in D. Cocultures were performed in three biological replicates and three technical replicates in B, C, and F. Experiments were conducted in four biological replicates in E. Statistical significance was determined in B through two-way ANOVA with Tukey’s multiple comparisons (*P ≤ 0.05, ***P ≤ 0.001); and in C with one-way ANOVA with Tukey’s multiple comparisons tests in GraphPad Prism (*P ≤ 0.05, **P ≤ 0.01).
Views: 125