

AI-designed OpenCRISPR-1 performs robust knockout, base editing, and prime editing in rice
Priya Das, Romio Saha, Debasmita Panda, Chandana Ghosh, S. P. Avinash, Sonali Panda, Mirza J. Baig, Kutubuddin A. Molla
New Physiolist; First published: 21 May 2026; https://doi.org/10.1111/nph.71272
ABSTRACT
RNA-guided programmable nucleases enable high-precision genome engineering for applications in agriculture, medicine, and biotechnology. Genome editing in biological organisms relies on a limited set of naturally occurring enzymes. Widely used nucleases such as Cas9 (Jinek et al., 2012) and Cas12a (Zetsche et al., 2015), as well as emerging effectors including TnpB (Karvelis et al., 2021; Karmakar et al., 2024a) and IscB (Altae-Tran et al., 2021), are derived from bacterial systems and often exhibit constraints in non-native eukaryotic contexts (Pacesa et al., 2024). Artificial intelligence (AI)-driven protein designing is now transforming nuclease engineering by leveraging large language models trained on the vast natural diversity of protein sequences (Nijkamp et al., 2023). Recently, OpenCRISPR-1 (OC1), an AI-generated RNA-guided nuclease comprising 1380 amino acids and differing from the prototypical SpCas9 by 403 amino acid substitutions, was shown to support efficient genome editing in human cells (Ruffolo et al., 2025). OC1 was released as an open-source nuclease and has been proposed to be freely available for licensing for academic research and commercial development (Ruffolo et al., 2025); consequently, it has the potential to substantially enhance global access to genome-editing technologies and represents an attractive alternative to the widely used SpCas9. In summary, we developed POC1-mediated versatile genome-editing platforms and demonstrated efficient NHEJ-driven indel formation, adenine and cytosine base editing, and prime editing in rice. Our results establish that the AI-designed nuclease OpenCRISPR-1 (Ruffolo et al., 2025), following codon optimization, is compatible with multiple genome-editing modalities and achieves performance comparable to that of the naturally occurring SpCas9 across diverse target loci in rice. Although the OC1 exhibited significantly reduced off-target editing compared to SpCas9 in human cells (Ruffolo et al., 2025), its off-target activity in plant systems remains to be evaluated. Similarly, the performance of OC1 should be assessed across a broader range of monocot and dicot species to support its wider adoption as an alternative to SpCas9. The expanding repertoire of AI-designed effectors (Thomson et al., 2025), including nucleases and deaminases, is poised to unlock new opportunities in eukaryotic genome engineering by overcoming intrinsic limitations of naturally evolved systems.
See https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.71272
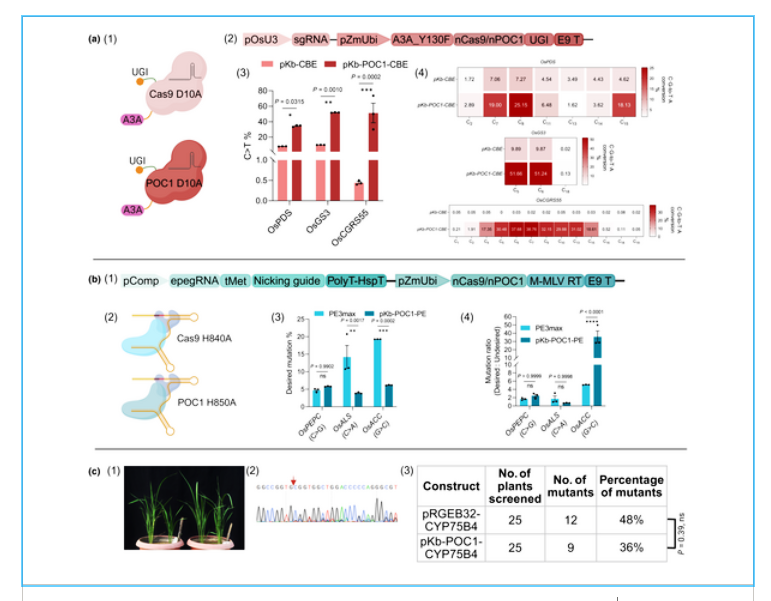
Figure 2
Plant OpenCRISPR-1 (POC1) mediates cytosine base editing, prime editing, and the generation of stable mutant lines in rice. (a1, a2) Schematic and cartoons of the pKb-CBE & pKb-POC1-CBE cassettes. (a3) Comparative C-to-T editing percentage. (a4) Editing window of CBEs at three targets. (b1, b2) Cartoons and schematic of the PE3max and pKb-POC1-PE cassettes. (b3) Efficiency of intended edits. (b4) Ratio of intended to unintended editing efficiencies. (c1) cyp75b4 knockout plant lines generated using POC1. (c2) Sanger chromatogram of a POC1-mediated cyp75b4 knockout plant showing the target locus with overlapping peaks indicative of biallelic heterozygous mutations. (c3) Comparison of editing efficiency in T0 plant lines for OsCYP75B4 target (ns, P > 0.05; *, P < 0.05; **, P < 0.01; ***, P < 0.001; ****, P < 0.0001). Error bars in the figures represent ± SEM. ns, non-significant. This figure was created in BioRender (BioRender.com/01an8ho).
Views: 11